The Gulf of Eilat-Aqaba is an oligotrophic sea (low in nutrients), located as the northeastern extension of the Red Sea. From the Gulf of Eilat and along the Red Sea (over 2000 km), there exists a gradient of increasing primary production and chlorophyll (Chlorophyll-a) values towards the source lake in the Arabian Sea (Figure 1). Primary production, the composition of phytoplankton, their distribution patterns, and seasonal dynamics in the Gulf of Eilat are influenced and change according to the seasonal cycles of stratification and vertical mixing in the water column (Genin et al., 1995; Labiosa et al., 2003; Shaked and Genin, 2023; Zarubin et al., 2017).
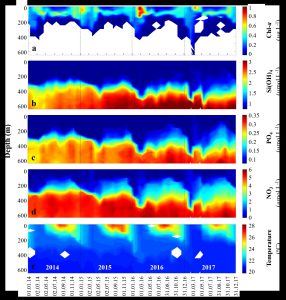
Figure 2. Oceanographic depth profiles from the surface to a depth of 600 m during the years 2014-2017: Chlorophyll concentrations (a), nutrient concentrations: silica (b), phosphate (c), nitrate (d), and temperature (e). The data were processed from results of the National Monitoring Program sampling (Shaked and Genin, 2017; https://iui-eilat.ac.il/Research/NMP). The figure is taken from Chernihovsky et al. (2020).
The periodicity of primary production varies seasonally in a distinct manner. During the summer months (April to September), the water column is stratified in a stable state, with a strong thermocline separating the upper layer from the deeper water layer. This stratification limits vertical mixing and nutrient exchange between the layers. The depth of the maximum chlorophyll concentration (Deep Chlorophyll Max) ranges between 80-100 m. The upper water layer is relatively warm (>25°C), nutrient-poor (NO3 ≈ 0 µmol/l, PO4 ≈ 0.06 µmol/l, Si(OH)4 ≈ 0.54 µmol/l), and contains chlorophyll concentrations of approximately 0.01 µg/l and primary production values of around 0.3 g C m-2 d-1. In contrast, the deeper water layer is colder (~20°C) and nutrient-rich (NO3 ≈ 6.7 µmol/l, PO4 ≈ 0.4 µmol/l, Si(OH)4 ≈ 3.5 µmol/l) (Figure 2, Shaked and Genin, 2023). During this period, the phytoplankton community is predominantly composed of picocyanobacteria of the Prochlorococcus spp. type (which make up about 40-50% of the total phytoplankton during this period) and Trichodesmium spp. algae (Lindell and Post, 1995; Post et al., 2002).
Towards the end of autumn and during the winter, a gradual decrease in surface temperature leads to deep vertical mixing (300-800 m) in the water column that lasts for several months (Genin et al., 1995; Shaked and Genin, 2023). The depth of vertical mixing is determined by the gradient of heat loss from the surface water to the atmosphere (Carlson et al., 2014). During vertical mixing, the deeper water layer mixes with the upper layer, enriching the illuminated zone (photic zone) with nutrients. As a result, primary production and chlorophyll concentrations increase (Levanon-Spanier et al., 1979; Labiosa et al., 2003; Lazar et al., 2008; Meeder et al., 2012; Zarubin et al., 2017). During this period, the ecosystem in the Gulf of Eilat is mesotrophic, and most of the phytoplankton community consists mainly of eukaryotic microalgae: dinoflagellates, diatoms, and coccolithophorids (Winter et al., 1979). Maximum primary production values recorded during this period reached 1.87 g C m-2 d-1 (Figure 2, Shaked and Genin, 2023). In the spring bloom, when chlorophyll concentrations peak (1.72 µg/l), the dominant phytoplankton are picocyanobacteria of the Synechococcus spp. type. These photosynthetic bacteria consume most of the remaining nutrients in the illuminated zone, causing the upper water column to become oligotrophic again in the summer months (Lindell and Post, 1995; Shaked and Genin, 2023).
Accordingly, the ecosystem in the Gulf of Eilat is primarily controlled by the availability of nutrients for primary producers—bottom-up control—rather than by grazing pressure from herbivorous organisms (top-down control) (Levanon-Spanier et al., 1979; Sommer, 2000).
References
Carlson DF, Fredj E, Gildor H (2014) The annual cycle of vertical mixing and restratification in the Northern Gulf of Eilat/Aqaba (Red Sea) based on high temporal and vertical resolution observations. Deep Sea Research Part I: Oceanographic Research Papers 84:1-17. DOI: 10.1016/j.dsr.2013.10.004.
Chernihovsky N, Torfstein A, Almogi-Labin A (2018) Seasonal flux patterns of planktonic foraminifera in a deep, oligotrophic, marginal sea: sediment trap time series from the Gulf of Aqaba, northern Red Sea. Deep-Sea Res. Part I: Oceanographic Res. Papers 140, 78–94. DOI: 10.1016/j.dsr.2018.08.003.
Chernihovsky N, Almogi-Labin A, Kienast SS, Torfstein A (2020) The daily resolved temperature dependence and structure of planktonic foraminifera blooms. Sci. Rep. 10, 17456. DOI: 10.1038/s41598-020-74342-z.
Genin A, Lazar B, Brenner S (1995) Vertical mixing and coral death in the Red Sea following the eruption of Mount Pinatubo. Nature 377(6549):507-510. DOI: 10.1038/377507a0.
Labiosa RG, Arrigo KR, Genin A, Monismith SG, van Dijken G (2003) The interplay between upwelling and deep convective mixing in determining the seasonal phytoplankton dynamics in the Gulf of Aqaba: Evidence from SeaWiFS and MODIS. Limnology and oceanography 48(6):2355-2368. DOI: 10.4319/lo.2003.48.6.2355.
Lazar B et al. (2008) Recent environmental changes in the chemical-biological oceanography of the Gulf of Aqaba (Eilat). Aqaba-Eilat, the improbable gulf. Environment, biodiversity and preservation. Magnes Press, Jerusalem 49-61.
Levanon-Spanier I, Padan E, Reiss Z (1979) Primary production in a desert-enclosed sea- the Gulf of Elat (Aqaba), Red Sea. Deep Sea Research Part A. Oceanographic Research Papers 26(6):673-685. DOI: 10.1016/0198-0149(79)90040-2.
Lindell D, Post AF (1995) Ultraphytoplankton succession is triggered by deep winter mixing in the Gulf of Aqaba (Eilat), Red Sea. Limnology and Oceanography 40(6):1130-1141. DOI: 10.4319/lo.1995.40.6.1130.
Meeder E, Mackey KR, Paytan A, Shaked Y, Iluz D, Stambler N, Rivlin T, Post AF, Lazar B (2012) Nitrite dynamics in the open ocean – clues from seasonal and diurnal variations. Marine Ecology Progress Series 453. DOI: 10.3354/meps09525.
Post AF, Dedej Z, Gottlieb H, Thomas DN, El-Absawi M, El-Naggar A, El-Gharabawi M, Sommer U (2002) Spatial and temporal distribution of Trichodesmium spp. in the stratified Gulf of Aqaba, Red Sea. Mar. Ecol. Progress. Ser. 239:241–250.
Sommer U (2000) Scarcity of medium-sized phytoplankton in the northern Red Sea explained by strong bottom-up and weak top-down control. Marine Ecology Progress Series 19-25. DOI: 10.3354/MEPS197019.
Shaked Y, Genin A (2017) The Israel National Monitoring Program in the Northern Gulf of Eilat/Aqaba. Scientific Report 2016. 181 p. http://www.iui-eilat.ac.il/Research/NMPMeteoData.aspx.
Shaked Y, Genin, A (2023) The Israel National Monitoring Program in the Northern Gulf of Eilat/Aqaba. Scientific Report 2022. 211 p. http://www.iui-eilat.ac.il/Research/NMPMeteoData.aspx.
Winter A, Reiss Z, Luz B (1979) Distribution of living coccolithophorid assemblages in the Gulf of Elat (‘Aqaba). Marine Micropaleontology 4:197-223. DOI: 10.1016/0377-8398(79)90017-3.
Zarubin M, Lindemann Y, Genin A (2017) The dispersion-confinement mechanism: Phytoplankton dynamics and the spring bloom in a deeply-mixing subtropical sea. Progress in Oceanography 155:13-27.