חברות הפיטופלנקטון והזואופלנקטון המגוונות במפרץ אילת-עקבה מציגות שינויים בתפוצתן המרחבית ובדינמיקה העונתית בהתאם לתנאים אוקיאנוגרפיים משתנים הכוללים: טמפרטורת מי הים, יציבות עמודת המים (שיכוב לעומת ערבוב עמודת המים) וזמינות נוטריינטים – חומרי תשמורת (Lindell & Post, 1995; Farstey et al., 2002; Chernihovsky et al., 2018).
במפרץ אילת מתועדים כ- 138 מיני פיטופלנקטון, כאשר הקבוצות העיקריות הינן: אצות כחוליות (בעיקר Prochlorococcus spp., Synechococcus spp. ו- Trichodesmium spp., Lindell & Post, 1995), אצות צורניות (80 מינים), דינופלג'לטים (57 מינים, Kimor & Golandsky, 1977; Winter et al., 1979; El-Sherbiny et al., 2019), וקוקוליטופורידים (52 מינים, Keuter et al., 2023). עושר המינים, המגוון הביולוגי והרכב המינים משתנים בהתאם לדינמיקה העונתית של עמודת המים ולזמינות הנוטריינטים (ראה פרק "יצרנות ראשונית, אצות וחומרי תשמורת").
חברת הזואופלנקטון במפרץ אילת מונה 30 טקסונים, כאשר הסרטנים מתת-מחלקת השטרגליים (Copepoda) הינם הדומיננטיים ביותר, כ-70 מינים מתועדים ובעלי שפעה בטווח בין 140 ל-517 פרטים למ"ק (El-Sherbiny et al., 2019). פירוט קבוצות האורגניזמים הזואופלנקטוניים הנוספים במפרץ אילת (ריסניות, ווליגרים של רכיכות, אצטלנים, תולעים וכו') מוצג בטבלה מס' 3 במחקרה של (2002) Farstey et al. , ובטבלה I במחקרם של (2019) . El-Sherbiny et al.
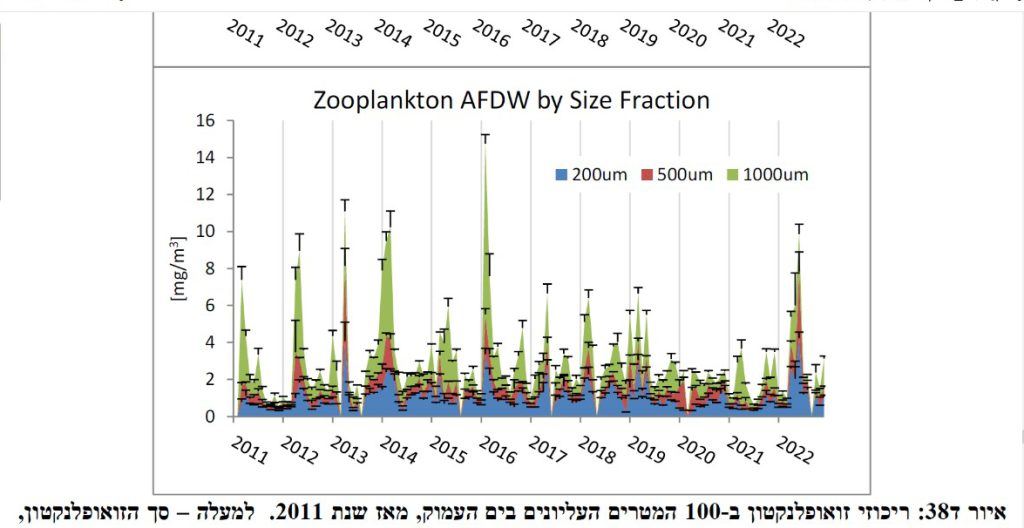
ביומסת הזואופלנקטון מציגה דינמיקה עונתית בהתאמה לזמינות מזונם המושפעת בעקיפין משינויי טמפ' פני השטח במפרץ אילת. במהלך הקיץ, עמודת המים העליונה משוכבת, ענייה בנוטריינטים, ומעל ל- 70% מהאורגניזמים הזואופלנקטוניים (100 < מיקרומטר) שוכנים באזור הפוטי המואר (100 מ'). לקראת סוף הסתיו ובמהלך החורף, ירידה הדרגתית של טמפ' פני השטח מובילה לערבול אנכי בעמודת המים של נוטריינטים משכבות המים העמוקות עם האזור המואר, דבר המעודד יצרנות ראשונית מוגברת, ריכוזי הכלורופיל מגיעים לשיאם ואף נצפית עלייה בריכוזי הזואופלנקטון. עם תחילת האביב, ביומסת האורגניזמים הזואופלנקטוניים מגיעה בשיאה עד כ- 24 מ"ג למ"ק (איור 1, Chernihovsky et al., 2018; Shaked & Genin, 2023). עומק הערבול האנכי במפרץ אילת הינו משמעותית עמוק (300-800 מ') ובתקופה זו כלל האצות והזואופלנקטון מפוזרים באופן הומוגני לאורך כל עומק הערבול(Lindell & Post, 1995; Kimor & Golandsky, 1977; Farstey et al., 2002; Keuter et al., 2023). למעט מספר מינים של סרטנים זואופלנקטוניים גדולים (750< מיקרומטר) כגון: Euphausiids, Isopods, Amphipods ו-Copepods אשר מבצעים נדידה אנכית אקטיבית, ובהתאם, התפלגותם האנכית שונה בין שעות היום לשעות הלילה (Farstey et al., 2002; El-Sherbiny et al., 2019).
המערכת האקולוגית במפרץ אילת כוללת רמות טרופיות רבות ונשלטת בעיקר על ידי זמינות נוטריינטים (bottom-up). בסביבות אוליגוטרופיות, לזואופלנקטון חשיבות רבה כחוליה מקשרת בין היצרנות הראשונית לרמות טרופיות גבוהות, המשפיעה על כלל מארג המזון הימי ועל הלולאה המיקרוביאלית (Levanon-Spanier et al., 1979; Sommer, 2000; Labiosa et al., 2003).
התחממות עתידית של טמפ' הים ושינויים בזמינות נוטריינטים, עשויים להוביל לשינויים בהרכב ובשפעת מיני הפלנקטון. שינויים אלו עלולים להשפיע על הרמות הטרופיות הגבוהות ובכך על המחזורים הביוגאוכימיים הימיים ובפרט על מחזור הפחמן (Frada et al., 2021).
איור 1 . ביומסה של זואופלנקטון (מ"ג למטר מעוקב) בעומק 0-100 מ' בשנים 2011-2022, בחלוקה לפרקציות גודל: 200-500 מיקרומטר (כחול), 500-1000 מיקרומטר (אדום) ומעל 1000 מיקרומטר (ירוק). נלקח מתוך Shaked & Genin (2023).
ביבליוגרפיה
Chernihovsky, N., Torfstein, A., & Almogi-Labin, A. (2018). Seasonal flux patterns of planktonic foraminifera in a deep, oligotrophic, marginal sea: Sediment trap time series from the Gulf of Aqaba, northern Red Sea. Deep Sea Research Part I: Oceanographic Research Papers, 140, 78-94.
El-Sherbiny, M. M., Devassy, R. P., Muller, E. M., Al-Sofyani, A. A., & Al-Aidaroos, A. M. (2019). Biodiversity and abundance of plankton communities along the coastal waters of Gulf of Aqaba, Saudi Arabia. Pakistan Journal of Zoology, 51(5), 1823.
Farstey, V., Lazar, B., & Genin, A. (2002). Expansion and homogeneity of the vertical distribution of zooplankton in a very deep mixed layer. Marine Ecology Progress Series, 238, 91-100.
Frada, M. J., Keuter, S., Koplovitz, G., & Avrahami, Y. (2022). Divergent fate of coccolithophores in a warming tropical ecosystem. Global Change Biology, 28(4), 1560-1568.
Keuter, S., Koplovitz, G., Torfstein, A., & Frada, M. J. (2023). Two-year seasonality (2017, 2018), export and long-term changes in coccolithophore communities in the subtropical ecosystem of the Gulf of Aqaba, Red Sea. Deep Sea Research Part I: Oceanographic Research Papers, 191, 103919.
Kimor, B., & Golandsky, B. (1977). Microplankton of the Gulf of Elat: aspects of seasonal and bathymetric distribution. Marine Biology, 42, 55-67.
Labiosa, R. G., Arrigo, K. R., Genin, A., Monismith, S. G., van Dijken, G. (2003). The interplay between upwelling and deep convective mixing in determining the seasonal phytoplankton dynamics in the Gulf of Aqaba: Evidence from SeaWiFS and MODIS. Limnology and oceanography 48(6):2355-2368. DOI: 10.4319/lo.2003.48.6.2355.
Levanon-Spanier, I., Padan, E., & Reiss, Z. (1979). Primary production in a desert-enclosed sea- the Gulf of Elat (Aqaba), Red Sea. Deep Sea Research Part A. Oceanographic Research Papers 26(6):673-685. DOI: 10.1016/0198-0149(79)90040-2.
Lindell, D., Post, A. F. (1995). Ultraphytoplankton succession is triggered by deep winter mixing in the Gulf of Aqaba (Eilat), Red Sea. Limnology and Oceanography 40(6):1130-1141. DOI: 10.4319/lo.1995.40.6.1130.
Sommer, U. (2000). Scarcity of medium-sized phytoplankton in the northern Red Sea explained by strong bottom-up and weak top-down control. Marine Ecology Progress Series 19-25. DOI: 10.3354/MEPS197019.
Shaked, Y., & Genin, A. (2023). The Israel National Monitoring Program in the Northern Gulf of Eilat/Aqaba. Scientific Report 2022. 211 p. http://www.iui-eilat.ac.il/Research/NMPMeteoData.aspx.
Winter A, Reiss Z, Luz B (1979) Distribution of living coccolithophorid assemblages in the Gulf of Elat (‘Aqaba). Marine Micropaleontology 4:197-223. DOI: 10.1016/0377-8398(79)90017-3.